基因的主宰:大脑中的信息与神经现实,以及自由意志

脑中的信息是什么?脑细胞如何制造与传递信息?大脑中的信息都储存在了哪里?大脑的可塑性又是什么?还有,大脑真的拥有自由意志吗?神经现实背后的主宰又是谁?
本文,将会从脑神经科学的角度——给出答案。
主题目录如下:
- 脑细胞不止一种
- 大脑里的信息是什么
- 电信息的制造
- 神经元的信息传递
- 突触的运作过程
- 神经元的可塑性
- 信息存储在了哪里
- 基因是主宰
- 那还有自由意志么
- One More Thing:随机试错
脑细胞不止一种
事实上,大脑中的细胞主要有两种:神经元细胞与神经胶质细胞。前者主要负责——处理和传递(与大脑功能相关的)信息;后者主要负责——结构支撑、保护绝缘、修复吞噬、以及营养供给等功能。
其中,神经胶质细胞主要(在大脑中的)有:
- 星形胶质细胞——结构支撑、平衡离子、吸收分子、营养供给、血脑屏障。
- 小胶质细胞——对抗感染、吞噬作用、免疫激活。
- 少突胶质细胞——保护绝缘、加速信息传递。
- 室管膜细胞——制造分泌脑脊液。
2009年,巴西神经科学家,苏珊娜·赫库拉诺·霍泽尔(Suzana Herculano Houzel )的论文估算出,大脑的神经元细胞约为860亿,非神经元细胞约为850亿——包括胶质细胞和其它细胞(如内皮细胞、上皮细胞)。
而我们通常所说的脑细胞,其实就是指神经元细胞,也简称为——神经元(或神经细胞)。
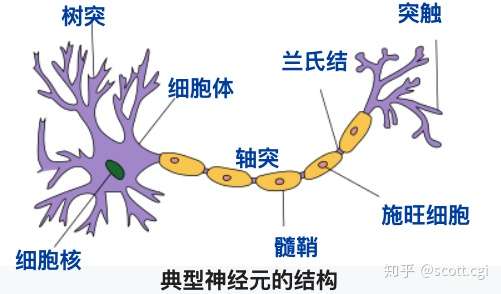
神经元(Neuron),由胞体(大而圆,Soma)和突起组成,突起又分为树突和轴突,连接在胞体上。
 图片来自维基百科
图片来自维基百科
- 树突(Dendrites)——多又短,反复分叉,由粗变细,呈树状分支。
- 轴突(Axon)——相对较长(最长可超过1米),有粗有细,直径均匀,只有一条。
- 髓鞘(Myelin)——呈节状相邻,长短不一(约60~1000μm),绝缘。
- 郎飞结(Nodes of Ranvier)——又称兰氏结,是髓鞘之间裸露的间隙。
- 轴丘(Axon Hillock)——是轴突起始的一小段(约15~25μm),无髓鞘部分。
- 神经纤维——是轴丘之后,由髓鞘包裹的部分**。**
- 神经末梢——是轴突末梢,分叉变细,呈树状分支的部分。
- 轴突末端(Axon Terminal)——是轴突末梢分支的末端。
- 突触(Synapse)——是轴突末端与靶细胞(其它神经元的胞体、树突、轴突)连接的结构。
极简形式如下示意:
靶细胞–突触—树突—胞体—轴突—突触–靶细胞
大脑里的信息是什么
神经元的核心功能之一,即是传递信息,那么这个可以“流动”的信息是什么呢?
显然,信息不能脱离物质独立存在,无论它是静止的还是流动的,它都需要载体。那么在大脑中,流动信息的物质载体——就是电与化学分子。因此,大脑中传递的信息——就是电信息与化学信息。
那电从何而来,电信息又是什么呢?
神经元的“电源”,主要来自细胞膜内外的带电离子,这些离子是:钠离子(正电)、钾离子(正电)、钙离子(正电)、氯离子(负电),以及有机负离子(Organic Anion,负电)。
离子——是指原子或原子基团,失去或得到一个或几个电子,而形成的带电荷的粒子。
我们知道,电荷分布形成了电位,分布差异形成了电位差。那么,膜内带电离子的流动就可以形成电位变化,而离子浓度就决定电位的强弱。
于是,电信息所传递的信息——就是电位的强弱变化,以及持续时间。
那化学信息是什么,又如何传递呢?
神经元可以制造化学分子,这是显而易见的,前面说的带电离子就是一种化学分子,但化学信息的传递,不是指带电离子,而是另外的一种化学分子——神经递质(后面具体说)。
那么,化学信息所传递的信息——其实就是神经递质的数量和类型。
于是,我们看到,化学分子——通过带电离子形成了电信息的传递,通过神经递质形成了化学信息的传递。
可见,大脑中信息的本质就是——化学分子,而分子是由原子构成的,原子是化学反应的最小单位(又称元素),再往后就是粒子物理了。
所以,大脑神经现实的运作,是在原子层面之上,以化学分子为基础的——这具有宏观确定性,而化学分子的运作**,是在原子层面之下(量子生物学),以量子效应**为基础的——这具有微观不确定性。
例如,在量子生物学中:
- 酶催的化反应,可以高速完成,可能是**“量子隧穿效应”**的体现。
- 光合作用的高效率,可能是**“量子相干性”**参与了能量的转移(否则,会存在随机的能量浪费)。
- 鸟类感知地磁场方向,可能是**“量子纠缠”**在起作用(鸟类视网膜内的自由基与电子纠缠)。
- 嗅觉的底层工作机制,可能是气味分子化学键的振动频率,产生了受体内部电子的**“量子隧穿效应”,**进而触发了嗅觉的神经信号传递。
电信息的制造
神经元制造电信息,有三种形式:静息电位、分段电位与动作电位。
电位,即电势;电位差,即相对电压。
第一,静息电位(Resting Potential)
顾名思义,就是在静息状态下(即没有接受任何信息时),神经元的细胞膜(包括胞体、树突、轴突),所具有的恒定电位差。此时,膜电位外正内负,如果把膜外电位看成0,那么膜内就大约是-60mV(负值表示膜内电位低)。
那为何静息状态下,会有电位差呢?
这是膜上的渗透通道(Leak Channel)与钠钾泵(离子传输器),动态调控膜内外离子浓度差,抵达扩散力与电动力平衡的结果。
- 扩散力——是离子浓度差带来的扩散作用。
- 电动力——是电荷相互吸引或相斥的作用,电位差会带来电动力,作用于膜内外的离子。
- 电化学驱动力——是扩散力与电动力的合力(净力)。
- 渗透通道——是永不关闭的,但不同的离子的通透性不同。
- 钠钾泵——是主动调控的通道,它会不断消耗能量(ATP),以进出2 : 3的比例,让钾离子进2,钠离子出3。
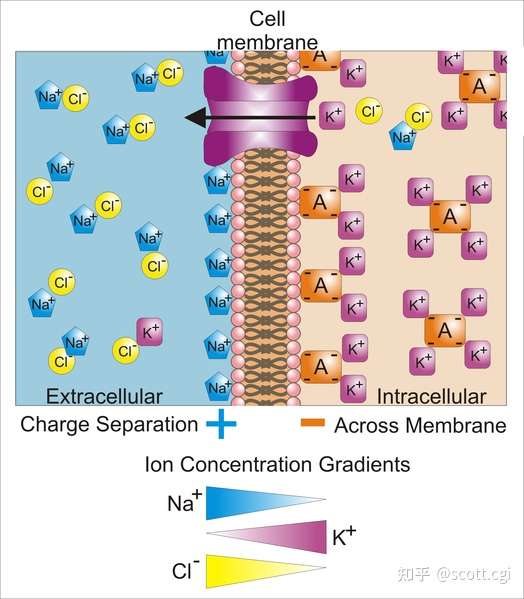 静息电位的膜电化学梯度,左边细胞外,右边细胞内,箭头表示库仑力持续抑制K+和Cl-的扩散,图片来自维基百科。
静息电位的膜电化学梯度,左边细胞外,右边细胞内,箭头表示库仑力持续抑制K+和Cl-的扩散,图片来自维基百科。
那么,静息膜内负电位的产生机制,就是:
- 钠钾泵——让膜内钾离子浓度升高,钠离子浓度降低,膜内负电位增强。
- 渗透通道——让高浓度钾离子扩散出去,膜内负电位增强。
- 渗透通道——准许氯离子进出,因为膜内负电位较强,电动力排斥氯离子出去,当膜外扩散力与膜内电动力平衡时,膜内负电位减弱。
- 渗透通道——不准许钠离子进出,无法改变电位差。
- 渗透通道——不准许钙离子进出,无法改变电位差。
- 有机负离子——没有通道准许进出,无法改变电位差。
我们可以看到,静息电位并没有传递信息,它只是维持了一个负电位状态,但这对神经元的信息传递至关重要。
第二,分段电位(Graded Potential)
这种电位是变化的、持续的、可叠加的、也是不断衰减的,并且只能在神经元的树突与胞体上触发传递。也就是说,分段电位是在从胞体与树突,向着轴突的轴丘,传递电信息。
其原理是,在胞体与树突上有一种特殊的通道,称为配体门控通道(Ligand Gated Channel)——它一直处在关闭状态,直到膜上受体与特定配体结合时,进入打开状态,此时通道可以准许一种或多种离子进入,这取决于通道的类型,而配体与受体分离时,通道就会关闭。
于是,我们就能看到:
- 当通道打开,特定离子进入膜内时,电位就会改变——这就是电信息的产生;
- 同时,这些离子聚集在狭窄的空间内,会有相斥的电动力和高浓度的扩散力,促使它们不断运动扩散——这就是电信息的传递;
- 但随着浓度下降,扩散速度会减慢——这就是电信息的衰减;
- 而如果通道一直打开,离子不断进入——这就是电信息的持续与叠加;
- 那么通道开闭断断续续,离子进入就会断断续续——这就是电信息的变化。
前面说过,电信息传递的就是——电位的强弱变化与持续时间,在此我们可以看到影响电信息传递的因素有如下:
- 其一,膜上配体门控通道的数量。
- 其二,靶细胞释放配体的数量。
- 其三,膜上受体的数量。
- 其四,配体与受体结合的时间。
而配体门控通道,主要有以下类型:
- 其一,钠钾钙离子-配体门控通道——由钠钾钙离子导致膜内,正电位增强。
- 其二,氯离子-配体门控通道——由氯离子导致膜内,负电位增强。
要知道,静息电位是负值(-60mV),那么“钠钾钙”带正电荷,就会减弱负电位的强度(如-50mV)——这被称为去极化(Depolarization),而“氯”带负电荷,就会增强负电位的强度(如-70mV)——这被称为超极化(HyperPolarization)。
这个意义就在于,当电信息抵达轴丘,如果其电位抵达了——阈电位(-50mV,Threshold Potential),就会触发轴突神经纤维上的电信息传递,否则将不会有信息传递。
因此,去极化会激发后续信息传递,我们称之为——兴奋电位;超极化会阻止后续信息传递,我们称之为——抑制电位;而轴丘,其作用也就是一个——触发区(Trigger Zone)。
最后,我们可以看到,分段电位不是恒定的,而是持续波动且可叠加的,这个叠加意味着,可以叠加增强,也可以叠加减弱——如同时开启**“正负电离子-配体门控通道”**。
第三,动作电位(Action Potential )
这个电位,就是在轴突神经纤维上,传递电信息的。其特点是,全无或全有,如果有就会以恒定的速度在神经纤维上,跳跃式传导,即:在无髓鞘的间隙速度稍慢,在有髓鞘的部分速度较快,但整体速度依旧很快,可以抵达每秒1~100米之间。
其原理是,在轴突上有一种特殊的通道,称为电压门控通道(Voltage Gated Channel)——它可以感应电位强度,以决定是否开启特定离子的通透性。
那么,在轴丘触发区的膜上,就聚集了大量的**“钠离子-电压门控通道”,一旦分段电位**在此处抵达阈电位(-50mV),通道就会迅速打开,接着大量钠离子涌入,电位就会一下飙升(到+40mV),那么高浓度的钠离子,在扩散力与电斥力的作用下,迅速流动,形成电信息的传递。
此时,“钠离子-电压门控通道”会因为电位远远偏离阈电位而关闭,产生绝对不应期(Refractory Period)即:无论什么强度的电位,都无法打开通道。
同时,另外一种**“钾离子-电压门控通道”,会稍慢一些感应打开,它会将膜内大量的钾离子排出,以及少量的“钾离子-渗透通道**”也会排除钾离子,从而降低膜内正电位,这一过程会使电位超极化(到-70mV)。
那么,当**“钠离子-电压门控通道”恢复后,此区域仍处在超极化状态,直到恢复到静息电位**(-50mV)之前——称之为相对不应期。因为,此时只能响应有限的电信息——需要有中和超极化的电位强度。
而在触发区之后,钠离子会继续扩散,从而形成**动作电位,**在神经纤维上快速传递,直到抵达神经末梢。
值得说明的是,在神经纤维上,因为有髓鞘覆盖,所以降低了膜外电位对内的影响,而髓鞘的间隙就会影响膜内电位,但这个间隙处会有**“钠离子-电压门控通道”,以增加钠离子浓度,从而抵消动作电位**的传递衰减,即化学扩散力的减弱。
因此,我们可以看到,在髓鞘处的电位传递速度,明显快于间隙处,呈现跳跃式传递。而不应期的存在,则阻止了动作电位的叠加与逆向。正是这些机制,保证了动作电位,可以高速稳定的传递电信息。
综上可见
电信息的制造机制,就是带电离子的浓度差,形成电化学驱动力(即扩散力与电动力的合力)驱动离子流动,从而产生了电位变化。
而神经元对离子浓度的控制,就在于膜上的各种离子通道,如:渗透通道、配体门控通道、电压门控通道、钠钾泵等等。
现在,我们就可以明白,为什么树突与胞体上的**“钠钾泵”,会消耗能量并源源不断地“出钠进钾”——因为动作电位的传递,是依靠逆向的“出钾进钠”**来实现的。
于是,树突与胞体就像是一个,电位的制造与控制器,其持续运作,产生可叠加的分段电位,一旦抵达阈电位,就会触发一次动作电位。
而分段电位有兴奋与抑制(即去极化与超极化),就是对触发动作电位的控制。也因此,动作电位只有兴奋,没有抑制——它的要的是效率,不是控制,其后续有化学信息的控制机制。
那么在整体上,膜内外就形成了一个,依靠能量运转的钠钾循环,其过程的目的和产物——就是制造和传递电信息。
概括起来,神经元的激活流程,即是:
神经元激活前,细胞膜处于静息电位(极化状态,外正内负,-60mV),当刺激引起膜外的正离子内流时,细胞膜发生去极化(少于-60mV),直到去极化抵达阈电位(-50mV)时,神经元激活,触发动作电位传递电信息(兴奋电位) ,此时**“钠离子-电压门控通道”打开导致大量钠离子内流,直到膜内电位达到绝对不应期**(+40mV),此时**“钾离子-电压门控通道”打开导致钾离子外流,直到膜内电位达到相对不应期**,发生超极化(-70mV,抑制电位),直到膜电位恢复到静息电位(-60mV),神经元激活结束。
最后,我们知道电荷的定向移动,产生了电流,那么神经元的膜内有电流存在吗?
显然,带电离子的扩散流动,虽然其过程充满了随机性,但整体上应该会是产生微弱的电流的。不过这个电流并不传递电信息,因为控制离子通道开闭的是电位差,而有离子流动,不一定就有电位差。
例如,持续相同强度的刺激,产生了膜内稳定的离子流动,但因为膜内外的离子浓度(在持续刺激下)趋于一致,无法产生**电位差,**所以就无法传递(刚开始刺激时的)电信息。
这就像令人快乐的行为,如果短时间内反复体验,就会无法让神经元产生电位差,从而失去最初的快感**,即“快乐的电信息”**。
由此,我们可以想象得到:
想法,并不是我们感觉上的抽象与虚无,因为每一个想法都是一个神经现实——是电化学反应在神经元之间电光石火般的碰撞,是一种信息的排列组合,是一种映射,是一种投影,是一种缩放的信息映射投影。
那么,一个想法的产生,就一定对应着这个想法的神经运作——从某种角度说,动作电位的传递就是想法的实体,离子通道的开闭就是想法的记忆。
神经元的信息传递
在大脑中,神经元自身的神经纤维,通过动作电位来传递电信息,但在神经元之间传递信息,却分为了两种情况,这依赖于神经元之间的连接方式,即:间隙中空(无物理接触)与间隙连接(有中空小桥接触)。
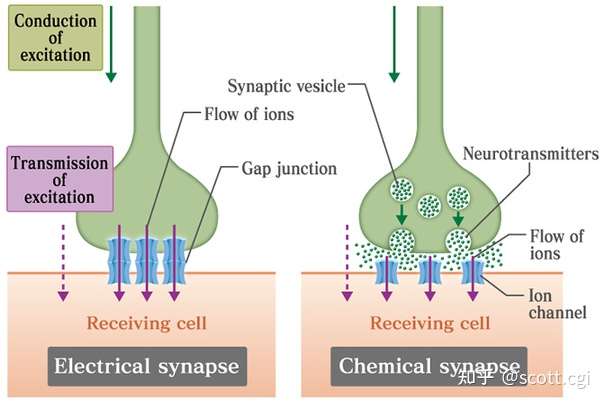 左边-电突触,右边-化学突触,图片来自必应搜图
左边-电突触,右边-化学突触,图片来自必应搜图
如果是间隙中空,这个结构被称为——化学突触,此时动作电位无法跨越间隙,于是电信息在此转化为化学物质,跨越间隙,在神经元之间形成化学信息的传递。
如果是间隙连接,这个结构被称为——电突触,此时动作电位可以直接通过(细窄的中空小桥),于是在神经元之间依然是电信息的传递。
例如,大脑皮层的星状细胞、小脑皮层的篮状细胞等,都有间隙连接,即有电突触。
由此可见,在神经元化学突触的连接路径上,其实是——电信息与化学信息交替接力传递的,即:神经元自身是电信息传递,神经元之间是化学信息传递。而神经元信息传递的过程,就叫做——神经脉冲(或神经冲动)。
那么,为什么神经元结构会在进化过程中,形成两种信息及混合传递的模式呢?
答案是——为了平衡,效率与控制。
电信息的传递效率——是显而易见的快,但过程难以控制,因为动作电位在传递中,要么不变,要么变小,无法增强。那么化学信息的传递,则就会更加的灵活,因为可以由化学物质的多少和类型,来动态控制电位传递的强弱。
这个化学物质,就是神经递质——可以称它为神经元之间的“信使”。
那么,动作电位的变化频率和持续时间,就可以在神经末梢,转换为神经递质的数量和类型,释放到化学突触,从而来控制后续神经元的活动状态。
而控制点就在于,可以动态的(根据具体环境和状态)决定——神经递质的运输、释放数量、与受体的结合时间,以及清除和回收速度,等等。
最后,结合信息传递,神经元表示成以下形式:
- 接收区(Receptive Zone)——树突和胞体。
- 触发区(Trigger Zone)——轴突的轴丘。
- 传导区(Conducting Zone)——轴突的神经纤维。
- 输出区(Output Zone)——轴突的神经末梢和突触。
即:–突触—树突—胞体—轴丘—神经纤维—神经末梢—突触–
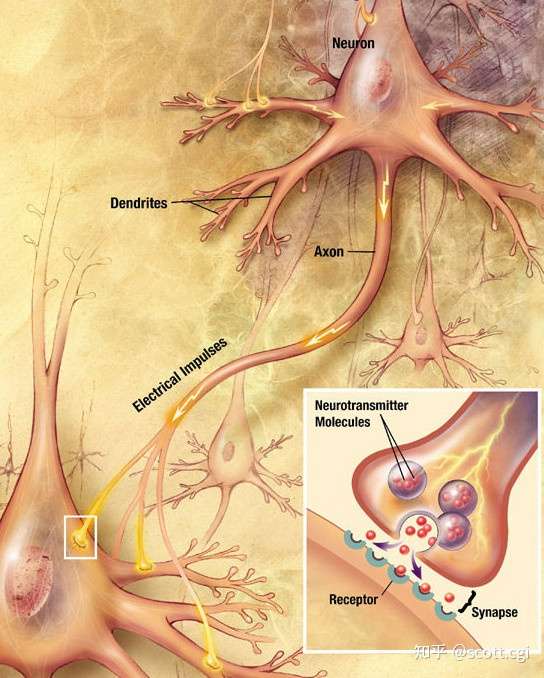
突触的运作过程
神经递质,会储存在突触小体中的突触小泡中。
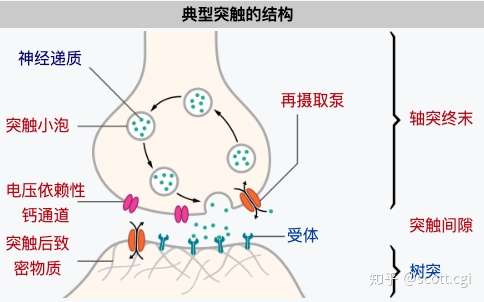 图片来自维基百科
图片来自维基百科
突触小泡 ——在神经元的不同部位形成,在小泡中含有高浓度的化学传递物质,随着神经末梢的兴奋,小泡的内含物向突触间隙释放,引起突触传递。
突触小体——是指轴突末端,膨大部分所形成的小体,它可以与其它神经元的胞体或树突相接触,形成突触。
当神经脉冲抵达神经末梢的时候,动作电位会改变突触前膜的膜电位,这会打开膜上的——钙离子-电压门控通道,于是钙离子就会进入突触小体,这又会激活一种酶,从而让突触小泡脱离细胞骨架,运动至突触前膜,接着在几种蛋白质的作用下发生膜融合,最后形成裂口通道,从而释放神经递质到突触间隙。
然后,神经递质就会扩散跨越间隙,涌向突触后膜,接着与膜上神经递质受体结合,从而打开特定的——配体门控通道,从而让特定的离子进入突触后膜,形成分段电位——激活后续神经元的电位变化。
于是,这就完成了一次——从电信息,到化学信息,再到电信息的传递——然后这个过程会在后续(由化学突触连接的)神经元之间循环发生。
而神经末梢一旦没有了动作电位(进入不应期),钙离子-电压门控通道就会关闭,从而钙离子浓度就会下降,突触小体就会停止释放神经递质,后续神经元的分段电位,也就会随即减弱,于是其轴丘电位,就无法抵达阈电位,也就无法触发动作电位。
那么,如果某一次的电信息过于强烈,释放了过多的神经递质到突触间隙,这时候突触前膜上,会有一种称为——再摄取泵的离子运输通道,将多余的神经递质运送会突出小体,以备后用。同时,突触间隙还会有一种酶,来分解过多的神经递质,防止后续神经元出现连锁性“过载”。
神经元的可塑性
事实上,化学信息的传递机制,化学突触的运作形式,就注定了神经元拥有——可塑性。
 图片来自维基百科
图片来自维基百科
那么,这个可塑性就在于:
如果,神经元经常接收到较强的信息(即被训练),那么基因就会表达出更多的神经递质(存于突触小泡),从而后续神经元,也就会接收到较强的信息,于是后续神经元的基因,就会表达出更多的神经递质受体,结果这条神经元的连接就被增强了——这代表着,神经元传递的信息,有更高的电位变化频率,即更强的神经脉冲。
相反,如果神经元经常接收到较弱的信息(即得不到训练),那么基因就会表达出更少的神经递质,接着后续神经元的基因,就会表达出更少的神经递质受体,结果这条神经元的连接就被减弱了——这代表着,神经元传递的信息,有更低的电位变化频率,即更弱的神经脉冲。
更甚的是,如果神经元的某个连接长期不“训练”,就会在基因的调控下,动态地断开连接,以腾出资源去强化更有效的连接路径。
而在整体的结构上,神经元之间的动作电位越强,基因就会表达出更多——树突与轴突的分支末端,以及突触连接。
那么,神经元的连接就会更多更密集,即:得到更为复杂的神经网络结构,显然这就是更强脑力的关键所在。
信息存储在了哪里
我们一直在讨论,神经元的信息传递,或许会有这样一个疑问——那就是这些信息会存储在神经元的什么地方呢?
答案就是——存储在了神经网络结构之中,即:结构即是信息。
更具体一些,信息是以神经连接形成的网络结构来存储的,即:连接结构编码了信息。
例如,看到一只从未见过的蝴蝶,其颜色信息会被视觉细胞捕获,其感知的结果是电脉冲(即动作电位),不同颜色的电脉冲抵达神经元,就会激活它们,进而向其它神经元,继续传递电脉冲,但这种电传递,会随距离及强度(幂律)衰减直至消失(即无法再激活神经元)——而这段时间内激活的神经元,所构成的**“路径结构”**,就是这只蝴蝶的信息。
事实上,神经网络是一个异常复杂的结构——大脑中约有860亿个神经元,其中每一个神经元,通常都与周围的神经元之间,平均约有7000个连接,可以想象整个网络结构的复杂度,就如同宇宙中的亿万星辰一般。
维基百科神经元: 成年人约有100~500万亿个突触连接
而神经元——既是信息传递和形成的单位结构,也是信息存储和处理的单位结构。神经元之间的几何关系、密度、数量,膜内外的成分、浓度、电位,以及电化学反应的过程,等等——都是一种信息的记录和计算。
那么,神经网络结构,可以动态的修改,即可塑性,就是信息的存储与清除——这对应着记忆与遗忘。
例如,每次获取相同信息,都在激活相同的路径结构,即记忆;长时间不激活,路径结构(其连接)就会减弱直至消失,即遗忘;获取与回忆激活的路径结构相似但不同,即偏差。
而类比来看,这个动态网络结构,就像是河床,其中的电化学反应,就像是河流——河床被河流塑造,即是结构被电化学反应的重塑,但同时河床也在引导河流的走向,即是结构在约束电化学反应的进程。
显然,这条脑内的“河流”,其蜿蜒的源头,来自于——环境信息。
如此,每个想法、观点、闪念,等等——都是大脑中神经脉冲的电位表达。而这一切,都是环境信息,涌入脑内结构之后发生的——河床与河流的引导与塑造。
最后,迈克斯·泰格马克在**《生命3.0》**一书中,进行了如下的估算:
- 人脑所有神经元,突触所能够存储的化学信息,大约是100T——这相当于硬盘。
- 人脑所有神经元,激活状态所能够存储的电信息,大约是10G——这相当于内存。
这个10G不知道是如何估算的,但这个100T是一个非常简单粗暴的估算——假设一个突触算1个byte,一个神经元有1000个突触连接,大脑算1000亿个神经元,于是有:
1000亿(神经元) * 1000(突触) = 1,000,000亿byte = 1000亿K = 1亿M = 100,000,000M = 100,000G = 100T。
那么,计算机与人脑在结构功能上的一个重要区别,就在于:
- 前者(冯·诺依曼构架)的信息存储与处理,在物理上是分离的,处理器与存储器之间需要进行大量的信息通信。
- 后者(人脑神经网络结构)的信息存储与处理,整合在了神经元的突触连接上,通过突触连接强度(即突触权重),实现了结构的可塑性,也实现了存储与处理的一体化,相比计算机,更加节能高效。
基因是主宰
如果说,使用化学信息传递信息,是为了可以使用神经递质,来进行神经脉冲的“行为”控制,那么我们不禁再进一步追问——是谁来控制神经递质这个“信使”的呢?
其实,从前面就已经可以,看出一些端倪了——神经递质是由基因调控表达的。
神经元(其胞体)会通过基因表达,来制造特定的神经递质与神经递质受体,而神经递质跨越突触,激活后续神经元的过程,则需要若干由基因表达制造的酶与蛋白质的协作,来完成神经递质的运输、释放、结合、以及回收。
让我们理清一下关系:
- 酶——是具有高度特异性和高度催化效能的,蛋白质或RNA。
- 蛋白质——是生命的物质基础,是有机大分子,是构成细胞的基本有机物,是生命活动的主要承担者。人体内蛋白质的种类很多,性质、功能各异。
- RNA——能够根据DNA序列,转运氨基酸、形成蛋白质模板、组成核糖参与合成蛋白质、以及成为特定的酶。
- 氨基酸——仅有二十几种,除了构成蛋白质,还能形成酸、激素、抗体、肌酸等含氨物质;转变为碳水化合物和脂肪;氧化成二氧化碳和水及尿素,产生能量。
- DNA——存储遗传信息的碱基序列,通常位于染色体上,非染色体上也有,比如线粒体与细胞质中。而人类体细胞染色体上的DNA共有碱基对三十亿个,其中绝大部分遗传信息并不会表达出来,同时对性状也没有直接的影响,这些是没有遗传效应的DNA片段,反之有遗传效应的DNA片段,就是基因。
- 基因——是具有遗传效应的DNA片段或RNA片段,即多个DNA或RNA,由碱基序列组成。基因编码了氨基酸和RNA,氨基酸组合形成肽链,多个肽链形成蛋白质。
综上来看,人体内的一切,其实都是——通过基因的编码,来翻译表达构造的。当然,大脑和大脑中的一切,包括神经现实的全部运作,也都是基因的杰作。
事实上,基因就是操控神经递质,来对神经系统施加直接影响的,而神经递质——就是神经现实运作中,形成各种宏观感受的核心所在,甚至(单胺类递质,如多巴胺、血清素、去甲肾上腺素等)还承载了意识、认知、思考、及注意力的底层构建。
可见,控制构建生命活动的信息,其实并不被我们主观意识所控制,甚至正是这些信息形成了我们的主观意识,而承载这些信息的就是基因。
是的,基因才是主宰,即:基因是主要的策略制定者,而大脑只是一个执行者。
那还有自由意志么
意识与智能、情绪与感受,是如此的复杂,以至于让人很难相信——其底层构建仅仅是:钠离子、钾离子、钙离子、电位变化、离子通道、化学分子、扩散力、电动力等等,这些简单到仿佛只有随机,而没有任何设计的物质与作用。
然而,就是随机试错,进化出了基因,以及上层所构建的生命。
事实上,我们所认为的“复杂”,包括这个“认为”本身的功能,就是基因利用神经递质对动作电位作出反馈,并进行了几十亿年随机试错的结果——显然,其中蕴含了“深邃”的策略、模式、以及极其复杂的网络协作性。
毫无疑问,几十亿年的随机试错——可以打败所有的智能与逻辑推理,这就是为什么到现在,我们依然还无法完全理解,大脑的功能与基因的算法。
那么说到自由意志,基因构建神经现实的目的只有一个——那就是对环境信息做出“最适合”的反馈,而自由意志就是这个反馈的——“最适合”。
其中,反馈是“半自由”的,因为神经现实具有可塑性,河流与河床可以相互影响,但终究“河床”以及一切,都会被“河流”的那个源头所影响和掌控——那就是环境信息,因此基因通过反馈给予的“半自由”,也必然成为了**“环境信息”手中的那根“提线”,成为了自我意识中的一种幻觉**,即是自由意志。
显然,幻觉是复杂的副产物(不是负),而如果大脑简单到,我们能够真正理解它(即没有幻觉),那么它就会过于简单,以至于我们没有足够的脑力,来做到理解它。因此,对于我们的脑力来说,大脑必然是复杂又充满了幻觉的。
是的,结论就是——人类根本没有所谓的自由意志,而只有集体的幻觉,即:虚幻的“自由感”。
智能与意识、喜好与情绪,都会随着病毒钻入神经元细胞,反转录其DNA而烟消云散。破裂的细胞,闪烁着线粒体最后的微光,照亮了正在一点点逝去的,凝固在DNA双螺旋结构里的灵魂。
One More Thing:随机试错
随机,在漫长的时间尺度下就会呈现出——规律,而在规律视角下,就拥有了方向,随机就变成了迭代——从而随机试错(演化),就变成了迭代试错(进化)。
Evolution——原本译作进化,后来发现进化带有目的性,大自然用演化更适合,但据我更细致的理解区分是,微观随机试错用演化,宏观迭代试错用进化。
是的,我们自由意志,只是微观局部的自由,在宏观整体上只有——确定的迭代与方向。